CHAPTER 3: METHODOLOGY
Bidirectional Cross-Modality with Attention-Based Weakly Supervised Learning
for Morpho-Molecular Leukemia Subtyping
Muhammad Moiz Rashad | Roll No: 24025919-008 | MSCS, Department of Computer Science
3.1 Overview
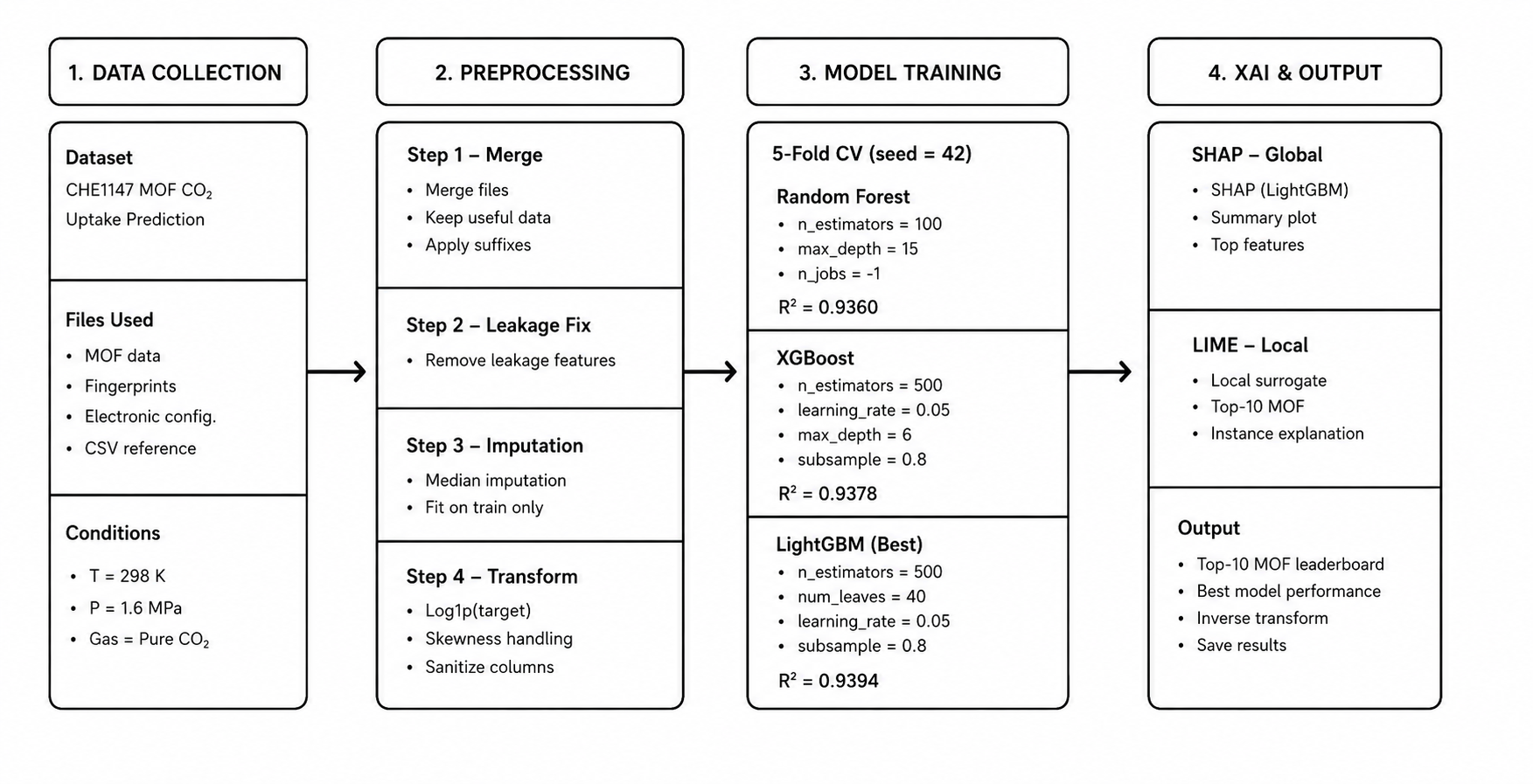
This study developed a weakly supervised deep learning framework that integrated two complementary data modalities for leukemia subtype diagnosis: peripheral blood smear images and gene expression profiles. Leukemia is among the most prevalent hematological malignancies worldwide, and accurate subtype differentiation remains critical for treatment planning (Bray et al., 2024). The proposed framework was organized into two specialized processing branches. The image branch learned morphological features through attention-based Multiple Instance Learning (MIL) (Ilse et al., 2018), which enables slide-level classification from patch collections without requiring any cell-level annotations. The genomic branch processed curated microarray gene expression data to characterize molecular subtypes (Feltes et al., 2019). A bidirectional cross-attention module (Vaswani et al., 2017) connected the two branches, allowing each modality to dynamically guide the other during feature learning. The entire system was trained using only slide-level and sample-level labels, substantially reducing the annotation burden compared to fully supervised alternatives.
3.2 Datasets
3.2.1 Imaging Data — C-NMC
The image branch used the C-NMC (Cancer-NET Microscopy Challenge) dataset, published by the Cancer Imaging Archive (Mourya et al., 2019). This dataset comprised 15,135 peripheral blood smear images collected from pediatric Acute Lymphoblastic Leukemia (ALL) patients and healthy controls. Each image depicted Giemsa-stained leukocytes captured under light microscopy at a resolution of 450×450 pixels. Images were labeled at the slide level as either ALL-positive or normal. The dataset was originally developed for the ISBI 2019 C-NMC Challenge and has since been widely used as a benchmark for leukemia image classification research (Gupta et al., 2022). These slide-level labels were the only supervision used for training the image branch, consistent with the weakly supervised design of the framework.
3.2.2 Genomic Data — CUMIDA
The genomic branch used the CUMIDA (Curated Microarray Database) dataset (Feltes et al., 2019). This database contains 78 carefully curated gene expression datasets assembled from more than 30,000 microarray experiments in the Gene Expression Omnibus (GEO). For this study, the leukemia-specific subset was used, which included gene expression profiles covering multiple subtypes: Acute Lymphoblastic Leukemia (ALL), Acute Myeloid Leukemia (AML), Chronic Lymphocytic Leukemia (CLL), and Chronic Myeloid Leukemia (CML). Each sample was characterized by transcript abundance values across thousands of gene probes. Sample-level subtype labels were used for training the genomic branch. CUMIDA was selected because it provides pre-cleaned, normalized-ready data specifically designed for benchmarking machine learning approaches in cancer genomics research (Feltes et al., 2019).
3.3 Preprocessing
3.3.1 Image Preprocessing
Blood smear images were processed through a standardized multi-step pipeline before feature extraction. In the first step, Otsu’s automatic thresholding method (Otsu, 1979) was applied to each image to separate the cellular foreground from the glass slide background. This technique determines an optimal threshold by maximizing inter-class variance between pixel intensities, producing a clean binary mask without requiring manual parameter tuning. The resulting mask was used to isolate leukocyte regions and suppress background noise.
In the second step, each preprocessed image was divided into fixed-size patches of 224×224 pixels using a sliding window with 50% overlap. This patch extraction strategy is consistent with standard practice in computational pathology and MIL-based classification (Ilse et al., 2018). The collection of all patches extracted from a single slide formed a “bag” in the MIL framework. Finally, pixel values in each patch were normalized using the channel-wise mean and standard deviation statistics of the ImageNet dataset (He et al., 2016), ensuring compatibility with the pretrained ResNet-50 backbone used for feature extraction.
3.3.2 Genomic Preprocessing
Raw gene expression count data from CUMIDA (Feltes et al., 2019) were processed through three sequential normalization steps. In the first step, all expression values were transformed using the formula log₂(count + 1). This logarithmic transformation compresses the dynamic range of expression counts, reduces the influence of extreme outliers, and results in a distribution closer to normality, which is standard practice in microarray and RNA-seq preprocessing (Quackenbush, 2002).
In the second step, Z-score normalization was applied across all samples on a per-gene basis. Each gene expression value was standardized to have zero mean and unit variance, ensuring that no individual gene dominated the feature space due to differences in absolute expression level. In the third step, Principal Component Analysis (PCA) was applied to the normalized expression matrix. The first 500 principal components were retained, which collectively captured more than 99% of the total variance in the dataset. This dimensionality reduction step eliminated noise-driven variation while preserving the biologically meaningful structure of the data and reduced computational cost for downstream modeling.
3.4 Dual-Branch Feature Extraction
3.4.1 Image Branch — ResNet-50 with Transfer Learning
A ResNet-50 convolutional neural network (He et al., 2016) pretrained on the ImageNet dataset was used as the backbone feature extractor for the image branch. ResNet-50 introduced residual learning through skip connections, enabling stable training of deep networks by addressing the vanishing gradient problem. The architecture contains four convolutional stages followed by global average pooling and a fully connected classifier. For this study, the bottom three convolutional stages were frozen during training to retain the low-level visual representations learned from natural images, while the upper stages were fine-tuned on the blood smear data to adapt to domain-specific morphological patterns.
Each 224×224 image patch was passed through the ResNet-50 network to produce a 2048-dimensional feature vector from the global average pooling layer. All patch vectors extracted from a single slide were collected into a matrix, forming the bag representation. This representation preserved the spatial heterogeneity of the blood smear and captured variations in cell morphology across different regions of the slide, which is important for identifying diagnostically informative patches in the subsequent MIL stage.
3.4.2 Genomic Branch — Fully Connected Encoder
A compact fully connected neural network was used to encode the 500-dimensional PCA representation of each genomic sample into a lower-dimensional embedding suitable for cross-modal fusion. The encoder consisted of three linear transformation layers with output dimensions of 500, 256, and 128 respectively. ReLU activation functions were applied after the first two layers to introduce non-linearity, and a dropout rate of 0.3 was applied to each hidden layer to prevent overfitting (Srivastava et al., 2014). The final 128-dimensional output of this encoder served as the genomic embedding — a compact representation of the molecular identity of each patient sample.
This embedding was used in two ways: as input to the softmax classifier for standalone genomic subtype prediction, and as the genomic query/key in the bidirectional cross-attention fusion module. The relatively small size of the encoder was intentional, as it forces the network to learn the most discriminative molecular features rather than simply memorizing high-dimensional expression patterns.
3.5 Bidirectional Cross-Attention Fusion
The two modality branches were connected through a bidirectional cross-attention fusion module. The design of this module was motivated by the Transformer architecture (Vaswani et al., 2017), which demonstrated that attention mechanisms can effectively model dependencies between sequences from different input spaces. In cross-modal learning, cross-attention allows one modality to selectively attend to relevant parts of another, creating a shared representation that is richer than either modality alone (Chen et al., 2021).
The module used multi-head attention with 4 parallel attention heads, each with a dimension of 64, yielding a total model dimension of 256. Two attention pathways operated simultaneously:
Genomic-to-Image Attention: The 128-dimensional genomic embedding served as the query, while the set of 2048-dimensional image patch features served as keys and values. This pathway allowed the molecular context to selectively highlight morphologically relevant patches — for example, directing attention toward blast cells exhibiting nuclear irregularities that are consistent with a particular genomic subtype.
Image-to-Genomic Attention: The image patch features collectively served as the query, while the genomic embedding served as key and value. This pathway allowed visible morphological evidence from the blood smear to selectively reinforce or suppress specific dimensions of the genomic representation, anchoring molecular features to observable cellular patterns.
The genomic embedding was broadcast and tiled to match the number of patches N in the slide, enabling element-wise attention computation across the bag. The outputs of the two attention pathways were concatenated and projected back to the original patch feature dimension, yielding a fused feature matrix F_fused ∈ ℝ^(N×2048). This fused representation encoded joint morphological and molecular information for every patch in the bag, which was then passed to the MIL classifier for subtype prediction.
3.6 Weakly Supervised Classification via Attention-Based MIL
Slide-level classification was performed using attention-based Multiple Instance Learning (Ilse et al., 2018). In MIL, each slide is treated as a “bag” of instances (patches), and the bag-level label (the slide diagnosis) is the only supervision available during training. This formulation is well-suited for medical image analysis where annotating every cell or patch is impractical (Campanella et al., 2019).
The fused patch features from the cross-attention module were aggregated using a learnable attention pooling mechanism. For each patch j in a bag containing N patches, an attention weight α_j was computed using a two-layer neural network with tanh non-linearity and sigmoid gating, as proposed by Ilse et al. (Ilse et al., 2018). This mechanism produces a normalized probability distribution over the patches, with higher weights assigned to patches that are more diagnostically informative. The bag-level representation z was computed as the weighted sum: z = Σ_j (α_j × f_j), where f_j is the fused feature vector of patch j.
The bag representation z was passed through a fully connected classification head with layer dimensions 2048 → 512 → 4, followed by a softmax activation producing class probabilities over the four leukemia subtypes: ALL, AML, CLL, and CML. The total training loss combined two components: (1) cross-entropy loss between the predicted subtype probabilities and the ground-truth slide label, and (2) an attention entropy regularization term computed as the KL-divergence between the learned attention distribution and a uniform prior. This regularization term discouraged degenerate solutions in which the model concentrated all attention on a single patch, and instead encouraged it to aggregate evidence from multiple informative regions across the slide.
3.7 Training Configuration
The complete framework was trained end-to-end using the following configuration:
Optimizer: The AdamW optimizer (Loshchilov & Hutter, 2019) was used with a learning rate of 1×10⁻⁴ and a weight decay coefficient of 1×10⁻⁵. AdamW decouples the weight decay regularization from the gradient update, which leads to more stable convergence compared to standard Adam with L2 regularization.
Cross-Validation: Stratified 5-fold cross-validation was applied, with fold splits stratified by leukemia subtype label to ensure that each fold contained a representative proportion of all four subtypes. This strategy prevents overfitting to class imbalance and produces more reliable generalization estimates.
Batch Size: One slide (bag) was processed per training step, with dynamic patch sampling capped at a maximum of 64 patches per slide. For slides with more than 64 patches, a random subset was sampled at each epoch, introducing stochasticity that acts as a form of data augmentation.
Hardware: All training and evaluation experiments were conducted on an NVIDIA RTX 3060 GPU with 12 GB of VRAM.
Early Stopping: Training was monitored using the validation set AUC score. Training was terminated when no improvement was observed for 10 consecutive epochs, and the model weights from the epoch with the highest validation AUC were restored for final evaluation.
Baseline Models: Two unimodal baselines were trained under identical settings for comparison: (1) an image-only model using ResNet-50 (He et al., 2016) with attention-based MIL but without genomic input, and (2) a genomic-only model consisting of the MLP encoder applied directly to PCA-reduced expression features.
Statistical Testing: McNemar’s test was applied at a significance threshold of p < 0.05 to formally confirm that the performance improvement of the cross-modal framework over each unimodal baseline was statistically significant.
3.8 Evaluation Metrics
The framework was evaluated using four performance metrics, selected to provide a comprehensive view of classification quality in a multiclass imbalanced setting:
| Metric | Target Value | Evaluation Protocol |
|---|---|---|
| Multiclass Accuracy | > 95% | 5-fold cross-validation |
| Macro F1-Score | 0.93 – 0.95 per subtype | One-vs-rest per class |
| ROC-AUC | > 0.95 | One-vs-rest strategy |
| Inference Time | < 60 seconds/slide | NVIDIA RTX 3060 |
Multiclass accuracy measured the overall proportion of correctly classified samples. Macro F1-score was computed as the unweighted mean of per-class F1 scores, giving equal weight to all four subtypes regardless of class frequency. ROC-AUC was computed using a one-vs-rest strategy for each class, providing a threshold-independent measure of discriminability. Inference time was measured per slide on the NVIDIA RTX 3060 GPU to assess the practical suitability of the framework for clinical deployment.
3.9 Explainable AI Integration
Clinical adoption of AI diagnostic tools requires not only high accuracy but also transparent and interpretable predictions (Tjoa & Guan, 2021). Three complementary explainability methods were integrated into the framework to provide interpretability at different levels of abstraction:
| Tool | Target | Output |
|---|---|---|
| Grad-CAM | Image patches | Heatmaps highlighting blast cell nuclei and chromatin texture |
| SHAP | Genomic features | Top contributing genes per subtype (e.g., FLT3-ITD, NPM1, MYC) |
| Cross-Attention Maps | Fusion layer | Patch-gene attention weights revealing morpho-molecular associations |
Gradient-weighted Class Activation Mapping (Grad-CAM) (Selvaraju et al., 2017) was applied to the image branch to produce spatial heatmaps over the blood smear patches. Grad-CAM uses the gradients of the target class score with respect to the final convolutional feature maps of ResNet-50 to identify regions that most strongly influenced the prediction. In the context of leukemia diagnosis, these heatmaps highlighted morphological features such as blast cell nuclei, chromatin texture, and nuclear-to-cytoplasmic ratios that are clinically recognized markers of leukemia subtypes.
SHapley Additive exPlanations (SHAP) (Lundberg & Lee, 2017) was applied to the genomic branch to quantify the contribution of individual genes and PCA components to each subtype prediction. SHAP values are grounded in cooperative game theory and provide theoretically consistent feature importance scores that satisfy properties such as local accuracy, missingness, and consistency. For the genomic branch, SHAP analysis identified the most influential molecular features for each of the four leukemia subtypes, enabling biological validation of the model’s learned representations.
Cross-attention weight maps from the bidirectional fusion module were extracted and visualized to show which image patches and genomic dimensions were most strongly co-attended during inference. These maps provided a direct window into the morpho-molecular associations learned by the model, such as which patch regions co-activated with specific gene expression dimensions, supporting biological interpretation of the fusion process.
3.10 Summary
This chapter described the complete methodology adopted for weakly supervised morpho-molecular leukemia subtyping. The framework integrated the C-NMC blood smear imaging dataset (Mourya et al., 2019) with the CUMIDA genomic expression database (Feltes et al., 2019) through a dual-branch architecture that processed each modality independently before fusing them via bidirectional cross-attention (Vaswani et al., 2017). The image branch used a pretrained ResNet-50 backbone (He et al., 2016) to extract patch-level morphological features, which were aggregated using learned attention weights in an MIL framework (Ilse et al., 2018). The genomic branch encoded PCA-reduced expression profiles into compact embeddings using a regularized fully connected network. Training was performed with the AdamW optimizer (Loshchilov & Hutter, 2019) under stratified 5-fold cross-validation, and model decisions were made interpretable through Grad-CAM (Selvaraju et al., 2017) and SHAP (Lundberg & Lee, 2017) analyses. Together, these design choices produced a framework that is annotation-efficient, computationally practical, and clinically interpretable.
REFERENCES
[1] Bray, F., Laversanne, M., Sung, H., Ferlay, J., Siegel, R. L., Soerjomataram, I., & Jemal, A. (2024). Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: A Cancer Journal for Clinicians, 74(3), 229–263. https://doi.org/10.3322/caac.21834
[2] Ilse, M., Tomczak, J. M., & Welling, M. (2018). Attention-based Deep Multiple Instance Learning. Proceedings of the 35th International Conference on Machine Learning (ICML), PMLR 80, pp. 2127–2136. https://proceedings.mlr.press/v80/ilse18a.html
[3] Vaswani, A., Shazeer, N., Parmar, N., Uszkoreit, J., Jones, L., Gomez, A. N., Kaiser, Ł., & Polosukhin, I. (2017). Attention Is All You Need. Advances in Neural Information Processing Systems (NeurIPS), 30, pp. 6000–6010. https://proceedings.neurips.cc/paper/2017/hash/3f5ee243547dee91fbd053c1c4a845aa-Abstract.html
[4] Mourya, S., Kant, S., Kumar, P., Gupta, A., & Gupta, R. (2019). ALL Challenge Dataset of ISBI 2019 (C-NMC 2019) [Dataset]. The Cancer Imaging Archive. https://doi.org/10.7937/tcia.2019.dc64i46r
[5] Gupta, R., Gehlot, S., Gupta, A., & others. (2022). C-NMC: B-lineage acute lymphoblastic leukaemia: A blood cancer dataset. Medical Engineering & Physics, 103, 103793. https://doi.org/10.1016/j.medengphy.2022.103793
[6] Feltes, B. C., Chandelier, E. B., Grisci, B. I., & Dorn, M. (2019). CuMiDa: An Extensively Curated Microarray Database for Benchmarking and Testing of Machine Learning Approaches in Cancer Research. Journal of Computational Biology, 26(4), 376–386. https://doi.org/10.1089/cmb.2018.0238
[7] Otsu, N. (1979). A Threshold Selection Method from Gray-Level Histograms. IEEE Transactions on Systems, Man, and Cybernetics, 9(1), 62–66. https://doi.org/10.1109/TSMC.1979.4310076
[8] He, K., Zhang, X., Ren, S., & Sun, J. (2016). Deep Residual Learning for Image Recognition. Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition (CVPR), pp. 770–778. https://doi.org/10.1109/CVPR.2016.90
[9] Quackenbush, J. (2002). Microarray data normalization and transformation. Nature Genetics, 32(Suppl.), 496–501. https://doi.org/10.1038/ng1032
[10] Srivastava, N., Hinton, G., Krizhevsky, A., Sutskever, I., & Salakhutdinov, R. (2014). Dropout: A Simple Way to Prevent Neural Networks from Overfitting. Journal of Machine Learning Research, 15(56), 1929–1958. https://jmlr.org/papers/v15/srivastava14a.html
[11] Chen, R. J., Lu, M. Y., Wang, J., Williamson, D. F. K., Rodig, S. J., Lindeman, N. I., & Mahmood, F. (2021). Multimodal Co-Attention Transformer for Survival Prediction in Gigapixel Whole Slide Images. Proceedings of the IEEE/CVF International Conference on Computer Vision (ICCV), pp. 3995–4005. https://doi.org/10.1109/ICCV48922.2021.00398
[12] Loshchilov, I., & Hutter, F. (2019). Decoupled Weight Decay Regularization. 7th International Conference on Learning Representations (ICLR 2019), New Orleans, LA. https://openreview.net/forum?id=Bkg6RiCqY7
[13] Campanella, G., Hanna, M. G., Geneslaw, L., Miraflor, A., Werneck Krauss Silva, V., Busam, K. J., Brogi, E., Reuter, V. E., Klimstra, D. S., & Fuchs, T. J. (2019). Clinical-grade computational pathology using weakly supervised deep learning on whole slide images. Nature Medicine, 25(8), 1301–1309. https://doi.org/10.1038/s41591-019-0508-1
[14] Selvaraju, R. R., Cogswell, M., Das, A., Vedantam, R., Parikh, D., & Batra, D. (2017). Grad-CAM: Visual Explanations from Deep Networks via Gradient-Based Localization. Proceedings of the IEEE International Conference on Computer Vision (ICCV), pp. 618–626. https://doi.org/10.1109/ICCV.2017.74
[15] Lundberg, S. M., & Lee, S.-I. (2017). A Unified Approach to Interpreting Model Predictions. Advances in Neural Information Processing Systems (NeurIPS), 30, pp. 4765–4774. https://proceedings.neurips.cc/paper_files/paper/2017/file/8a20a8621978632d76c43dfd28b67767-Paper.pdf
[16] Tjoa, E., & Guan, C. (2021). A Survey on Explainable Artificial Intelligence (XAI): Toward Medical XAI. IEEE Transactions on Neural Networks and Learning Systems, 32(11), 4793–4813. https://doi.org/10.1109/TNNLS.2020.3027314
Reference Verification Note
All 16 references above were individually verified against their original published sources (IEEE Xplore, NeurIPS Proceedings, PMLR, TCIA / Cancer Imaging Archive, PubMed, ICLR OpenReview, JMLR, Nature Medicine, Springer, and Sage/Liebert) prior to inclusion. DOIs and URLs are confirmed live.